
The genetic material in eukaryotic cells stays in a complex structure. This consists of DNA and proteins and is there in a specific compartment. It’s there as the nucleus. We know this structure as chromatin (from the Greek “chroma,” meaning coloured and “soma,” denoting body). About two meters of DNA must be turning into a tiny nucleus around a millimeter in diameter in each cell. Despite its extreme compactness, DNA must be easily accessible to interact with protein machinery. This controls chromatin functions.
Chromosomes in the interphase nucleus are difficult to distinguish from one another. Nonetheless, they include a distinct region within a core — a chromosomal area. On the other hand, lighter-stained euchromatin (transcriptionally active) and darker heterochromatin (transcriptionally quiet) are easily available. During cell division, chromosomal regions become very condensed chromosomes. We may distinguish them from one another. Mitotic chromosomes under a light microscope as a group is popular as karyotypes.
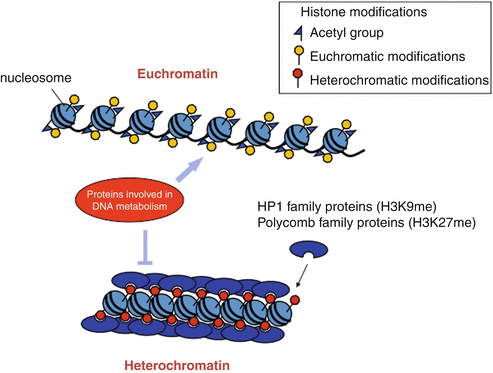
Heterochromatin and Euchromatin differences
The main differences between Heterochromatin and Euchromatin are:
- Euchromatin is in a free form, less vulnerable chromatin structure. At the same time, heterochromatin is in a dense packing. It is a very dense chromatin structure.
- GTG banding causes the euchromatin area to stain softly. But the heterochromatin region tarnishes darker. This explanation is that euchromatin’s loosely packed DNA absorbs less dye than the thicker heterochromatin area. The euchromatin is there mainly on the chromosome arms. Whereas heterochromatin is there primarily in centromeric and telomeric regions of the chromosomes.
- Histone proteins are crucial in determining which kind of chromatin forms during the DNA packaging process. In addition, the euchromatin zone fixes up or cannot spread. But the heterochromatin region can extend into the euchromatin region. This indicates that genes might become dormant as a result of a cell’s signal.
- Because genes are function bits of DNA that produce protein, nearly all are filled in the euchromatin area. However, this is not the case in the heterochromatin area.
- Because the heterochromatin is in a dense packing, the polymerase cannot replicate as quickly as it does in the euchromatin area. Hence, the euchromatin replicates first, followed by the heterochromatin.
- Genes are there in areas of euchromatin. We know this as ‘gene-rich regions.’ Whereas heterochromatin portions are ‘gene-less regions.’
- During the interphase of cell division, transcriptionally active chromatins are there on the inner side of the nucleus. Whereas inactive (heterochromatin) areas are located on the periphery of the heart.
- During metaphase, however, chromosomes contain both kinds of chromatins. They arrange themselves one after the other (therefore, we get light and dark bands simultaneously).
The lesser-known differences
- Only around 2% of the genome is genetically active or includes genes, remaining inert. Approximately 97–98% of the genome is simply trash, inactive heterochromatin.
- If we look at the chemical structure of both, we can see that the nature of nucleotide sequences differs. The euchromatin area is simple, with less repetitive DNA and GC-rich sections, whereas the heterochromatin region is highly redundant with GC-rich regions.

Heterochromatin definition
It is the genetically inactive part of the gene. Heterochromatin got its name from its nature to stain more heavily than most chromosomal ingredients. This happens throughout our cell cycle.
Types of Heterochromatin
There are two types of heterochromatin. The first one being constitutive HC and the later facultative HC. These differ slightly. This depends on the DNA that they contain. The richness in their satellite DNA determines the nature of the heterochromatin. This is polymorphism, and it has stain properties.
Constitutive heterochromatin
- Constitutive HC contains a form of satellite DNA. This creates many short, tandemly repeatable sequences: alpha-satellite DNA, DNA satellite I, II, and III. These satellite DNA sequences can fold on themselves. Thus they play a crucial role in developing the constitutive HC’s very compact structure.
- Constitutive HC is stable and retains its heterochromatic characteristics throughout development and in all tissues.
- Because of the fragility of the satellite DNA, constitutive HC is highly polymorphic. This polymorphism, which appears to have no phenotypic consequence. It can vary the size and the localization of the heterochromatin.
- The C-band method significantly stained constitutive HC due to the satellite DNA’s highly quick renaturation following denaturation.
Facultative heterochromatin
- The presence of LINE-type repeating sequences distinguishes facultative HC. These sequences, which are available across the genome, may encourage the spread of a condensed chromatin structure.
- Facultative HC is reversible. It has a heterochromatic status changing depending on the stage of development or cell type studied. Two instances of facultative HC are the inactive X (Barr body) in female somatic cells. It is also the inactive sex vesicle at the pachytene stage of male meiosis.
- Facultative HC has a low concentration of satellite DNA and so is not polymorphic.
- The C-band technique is not usable in staining facultative HC.

Properties of heterochromatin
Condensed heterochromatin
This is what heterochromatin is, and it applies to both constitutive and facultative HC. Because of the high condensation, it is highly chromophilic and inaccessible to DNAse 1 and other restriction enzymes in general.
Heterochromatin DNA replicates late.
The inclusion of different nucleotide analogs demonstrates that both constitutive and facultative HC DNA is late replicating. On the one hand, HC late replication is caused by its high condensation. This inhibits the replication machinery from quickly reaching the DNA. On the other hand, it is there for its placement in a peripheral nuclear domain deficient inactive elements.
DNA in heterochromatin is methylated.
On the cytosines, the DNA of constitutive HC is heavily methylated. As a result, an anti-5-methylcytosine antibody robustly identifies all constitutive HC areas.
In facultative HC, DNA methylation is more discrete. But the restriction enzymes are sensitive to methylation of CpG islands. They are preferentially available in gene regulatory areas.
Histones are hypo-acetylating in heterochromatin.
Histones’ N-terminal ends can undergo post-translational changes. This can alter the chromatin’s genetic activity. Hypo-acetylating of histone N-terminal tails, particularly on lysines, is in link with inactive chromatin. The active chromatin, on the other hand, is distinguishable as hyper-acetylated histones.
Read Also: Electric Force: Definition & Equation
Acetylation/deacetylation of histones is a critical process for controlling gene expression. Numerous transcription factors have been exhibiting Histone Acétyl Transférase or Histone Deacetylase activity.
Heterochromatin histones are methylation on lysine 9
Methylation of histone H3 lysine 9 (H3-K9) has only recently got involved in the process of genome heterochromatinization. This is both in constitutive and facultative HC.
Heterochromatin has a social instinct.
Various species’ studies have revealed that constitutive HC has a true propensity to collect during interphase. During interphase, the centromeres of polytene chromosomes, rich in heterochromatin, can combine to create centers in Drosophila larvae.
The number of heterochromatic blocks visible in interphase nuclei in the mouse is always less than the number of heterochromatic areas visible on metaphase chromosomes.

Euchromatin definition
Euchromatin is a kind of chromatin that is lightly packed (DNA, RNA, and protein) and is abundant in genes. It is frequently (but not always) under active transcription. Within the cell nucleus, euchromatin is the busiest part of the genome. The human genome contains 92 percent euchromatic DNA.
The structure of euchromatin is similar to an unfurled string of beads, with the dots representing nucleosomes. Nucleosomes are made up of eight proteins called histones and around 147 base pairs of DNA wrapped around them; in euchromatin, this wrapping is complimentary enough that the raw DNA may be accessible. Each core histone has a ‘tail’ structure that may vary in various ways; these variations are considered to serve as “master control switches,” determining the overall layout of the chromatin. It is thought that the presence of methylation lysine four on histone tails functions as a universal signal.
Euchromatin does not have any more types. It’s considered to be similar and unique in its way.
Properties of Euchromatin
Order of Evolution
In the case of the bacterial genome, euchromatin is the sole conformation of chromosomes. This implies that it evolved earlier than heterochromatin. Euchromatin, unlike heterochromatin, does not occur in two patterns. It exists exclusively as constitutive euchromatin. Euchromatin is necessary because it includes genes that are transcribed into RNA and subsequently into proteins.
Light Colored after Staining
Euchromatin is found in the nucleus’s core and accounts for around 90% of an organism’s DNA. After staining, it appears as light-coloured bands under an optical microscope. Heteropycnosis does not occur since all portions of euchromatin are coloured uniformly.
However, under an electron microscope, it appears as an elongated 10 nm microfibril.
Tightly and Uniquely Packed
The euchromatin format may be thought of as an unfurled string of beads representing nucleosomes. The nucleosomes house histone proteins, which coat a certain number of DNAs. Because the wrapping around by histone proteins is loosened in euchromatin, individual DNA sequences may be accessible. The shape of euchromatin is controlled by a methylation region of the chromosome known as the histone tail.
The sequence of Conversion
The unfolded shape of DNA in euchromatin allows regulatory proteins and RNA polymerase to connect to the sequences, allowing the transcription process to begin. Some genes in euchromatin can turn into heterochromatin. This is when they are no longer transcribable and are no longer active. The change of euchromatin to heterochromatin serves as a type of gene expression and replication regulation. Some genes, such as housekeeping genes, are always organizing euchromatin security for this reason. They must duplicate and transcribe constantly.



